Stephen K. Tyring, MD, PhD, MBA
Departments of Dermatology, Microbiology/Molecular Genetics and Internal Medicine, University of Texas Health Science Center, Houston, Texas
Disclosure: Dr. Tyring has conducted investigator-initiated clinical trials sponsored by PharmaDerm®, a division of Fougera Pharmaceuticals Inc.
Abstract
Background: Green tea catechins possess a wide range of pharmacological properties, including antiviral, anti-infective, and immunostimulatory properties. They also have demonstrated inhibitory effects on a variety of enzymatic and metabolic pathways involved in cancer development. Catechins have been shown to have antiproliferative properties in various cell lines and may have direct virucidal effect. The United States Food and Drug Administration has approved a topical ointment formulation of sinecatechins, derived from green tea catechins and other tea components, for the treatment of external genital and perianal warts. The exact mechanism of action of sinecatechins in eradication of human papillomavirus-induced external genital and perianal warts is unknown, but may be due to one or more of the mechanisms mentioned. Objective: This study was conducted to investigate the growth inhibitory potential of the sinecatechins in human cervical carcinoma cell lines infected with human papillomavirus. Methods: The viability of tumor cell lines (CaSki and SiHa infected with human papillomavirus-16; HeLa and C4-I infected with human papillomavirus-18) was investigated as one parameter in a short-term viability assay (48 hours). This was followed by a long-term clonogenic assay (12–23 days) to determine the cytotoxic potential of sinecatechins as a parameter for cell viability and proliferation. This assay determined if the effect observed in the viability assay was due to retardation in cell proliferation or to a reduction of total cell number, leading to cell death. Results: Based on the data collected, sinecatechins inhibited cell growth in all four tumor cell lines by 50 percent (GI50) at concentrations ranging from 160 to 360µM. C4-I cells were the most sensitive to treatment with sinecatechins, with a lower GI50 (~34µM). Total GI was achieved in a 48-hour assay at 625µM sinecatechins (40µM for C4-I), with growth inhibitory potential detectable after one hour. Clonogenic assays confirmed the cytotoxic potential of sinecatechins with a reduction in clone numbers in a concentration-dependent fashion. Sinecatechins substantially reduced the number of surviving HeLa cells at a concentration of 200µM, while surviving SiHa cells were almost totally eradicated with a concentration of 600µM. Conclusion: Sinecatechins demonstrated growth inhibitory potential in all four human papillomavirus-infected tumor cell lines, which may be attributed to the induction of apoptosis, mediated by cell cycle deregulation. In addition, this antiproliferative effect may contribute to the overall cancer-preventative function and possible direct antiviral activity of sinecatechins that may contribute to external genital and perianal warts clearance. (J Clin Aesthet Dermatol. 2012;5(2):34–41.)
The medicinal properties of tea leaves have been known for thousands of years.[1,2] Tea leaves, produced from the plant Camellia sinensis, contain two unique bioactive compounds—flavonoids and methylxanthines.[3] Methylxanthines, including caffeine, theobromine, and theophylline, are a group of alkaloids that are present in small quantities. Flavonols, which are a subtype of flavonoids, and also known as catechins, are the polyphenolic compounds that constitute 25 to 35 percent of green tea and are responsible for most of its beneficial effects.[2–4] Epigallocatechin-3-gallate (EGCg) is an important constituent of catechins and considered the predominant therapeutic agent derived from green tea.[3–5] Catechins, namely EGCg, have a wide variety of beneficial effects, including assisting in the prevention of chronic diseases ranging from cancer to cardiovascular diseases.[3]
Spectrum of biological and pharmacological activity of catechins. The spectrum of biological and pharmacological properties of green tea catechins includes antiangiogenic[4,6–13] activity, anti-inflammatory and immunostimulatory[2,3,5,8,10,11] activity, and antimicrobial potential.[14–22]
The antineoplastic activity of EGCg includes influencing tumor cell growth by inhibiting proliferation and inducing apoptosis.[3,4] EGCg has been shown to inhibit growth and induce apoptosis in head and neck squamous cell carcinoma,[6] and liver,[8] ovarian,[7] prostate,[9,10] breast,[11] and cervical cell lines.[12] Antiproliferative and anticarcinogenic effects of EGCg have also been demonstrated in vitro in lung cancer cells,[13] colon tumors, as well as lymphoma and leukemia cells.[4] Tumor reduction induced by EGCg has been demonstrated in breast cancer cell lines.[11] EGCg was shown to play an important role in inhibiting growth in a human papillomavirus (HPV) type 16-associated human cervical carcinoma cell line, CaSki. EGCg significantly inhibited growth by inducing apoptosis, possibly by negatively influencing cell proliferation at the G1 phase and gene expression pattern in CaSki cells.[4] Similar results were seen in human prostate cancer cells, where EGCg was shown to affect mitogenesis and induce apoptosis by causing G0/G1-phase cell cycle arrest.[9] Induction of apoptosis and cell cycle arrest at the G1 phase was also seen in ovarian and liver cancer cell lines.[7,8] In the androgen-sensitive human prostate carcinoma cells, LNCaP, EGCg was shown to induce apoptosis through stabilization of p53 by phosphorylation on critical serine residues, and by negative regulation of nuclear factor-kB (NF-kB) activity, a family of transcriptional regulators.[10] Upregulation of p53 expression was also seen in liver cancer cells,[8] and reduction in NF-kB levels and activity by inhibition of the Her-2/neu signaling pathway was seen in breast cancer cells.[11] Additionally, EGCg has also been associated with telomerase inhibition, a potentially selective target for treatment of cancer.[12] However, despite the various mechanisms investigated, including modulation of signal transduction pathways, regulation of gene expression, induction of apoptosis, inhibition of cell proliferation and transformation, the exact mechanism of action of EGCg in inhibiting neoplastic growth is still unclear.
EGCg also influences immunostimulation and anti-inflammatory responses by inducing the release of immune stimulatory agents while suppressing the release of immune inhibitory agents. The anti-inflammatory activity of EGCg may be related to an antioxidant function mediated by nonantioxidant mechanisms.[2,3] This includes the ability of EGCg to modulate signal transduction pathways8,10 and inhibit the activity of transcription factors, such as NF-kB[10,11] and activator protein 1 (AP-1).[2] The antioxidant function of EGCg is characterized by its ability to scavenge reactive oxygen species (ROS),[3,5] which also plays a key role in enhancing its anti-inflammatory properties.[5] For example, the downstream effect of inhibiting NF-kB activation results in downregulation of interleukin-8 (IL-8), a major neutrophil chemoattractant and inflammatory mediator.[2,5] In addition to IL-8, EGCg modulates inflammatory signaling pathways and negatively influences the infiltration of T cells into sites of inflammation.[5] Additionally, catechins activate T lymphocytes and induce the release of tumor necrosis factor alpha (TNF-a) and interferon-gamma (IFN-g). Immunostimulatory activity of catechins includes stimulating macrophages to release cytokines, promoting recruitment of monocytes, dendritic cells, lymphocytes, natural killer cells, and T-helper cells.[2]
EGCg has demonstrated a broad spectrum of antimicrobial activity against bacteria and fungi, including growth inhibition of Escherichia coli, Staphylococcus aureus, and Trichophyton sp.14,23 Most notable is the ability of catechins, such as EGCg, to protect against viral infections, including rotavirus,[14] enterovirus,[14,24] adenovirus,[16,24] human immunodeficiency virus (HIV),[17–19] Epstein-Barr virus (EBV),[14] and influenza A and B.[20–22]
Catechins have exhibited antivirulent activity against HIV through several steps in the HIV life cycle. EGCg not only prevents HIV attachment and cell entry, but also destroys viral particles and virus production by competitively inhibiting HIV-1 deoxyribonucleic acid (DNA) polymerase and reverse transcriptase. Viral transcription is also impacted with a distinct decrease in the expression of messenger ribonucleic acid (mRNA) in the presence of EGCg.[17,18] Furthermore, EGCg has been shown to inhibit HIV infection and replication without any observed apoptosis or necrosis.[19]
EGCg inhibits expression of EBV viral protein by inhibiting transcription of immediate-early genes, thus preventing initiation of EBV lytic cycle.[14] In Madin-Darby canine kidney (MDCK) cells, EGCg affected the infectivity of influenza A and B viruses by agglutinating the virus, thus blocking the attachment of the viral particles to the target cell receptors and preventing the virus from absorbing to MDCK cells.[20,21] In addition to preventing infection, EGCg was also shown to inhibit replication of the viruses by preventing acidification of endosomes and lysosomes, an essential step in uncoating of the virus and viral replication.[21,22]
Sinecatechins. Sinecatechins ointment, 15%, is manufactured from the extract of green tea leaves from Camellia sinensis, consisting of 85- to 95-percent catechins, with EGCg representing the primary catechin at more than 55 percent (Figure 1).[25] It is the first botanical agent approved by the United States Food and Drug Administration (FDA) for prescription use and is indicated for use as a topical patient-applied treatment for external genital and perianal warts (EGW).[26] Two Phase 3 clinical trials involving more than 1,000 male and female patients with EGW treated for up to 16 weeks with sinecatechins ointment, 15%, resulted in statistically superior complete clearance rates of all warts (baseline and newly emerging warts) compared with vehicle-treated patients (54.9% vs. 35.4%, respectively, p<0.001).[27] In addition, recurrence of warts was assessed in all patients with complete clearance of all warts; low rates of recurrence (6.8%) were demonstrated with sinecatechins ointment, 15%, when measured at 12 weeks post-treatment.[27] As opposed to previous clinical trials of EGW therapies, which only measured the clearance of baseline warts, clearance of all (baseline and new) warts represents a much more relevant therapeutic endpoint.[27]
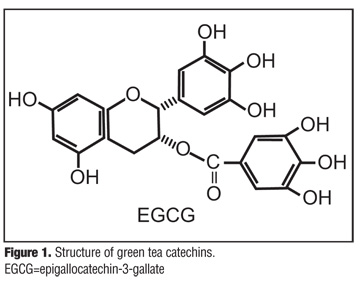{kind=link}
Although the molecular mechanisms of the antiviral and anticarcinogenic potential of green tea catechins have been elucidated recently in numerous studies, the mechanism of action (MOA) of sinecatechins in the eradication of EGW is unknown. As an anticarcinogenic agent, it has been shown that green tea catechins interfere with different biochemical pathways involved in tumor proliferation, mitogenic signal transduction, cell cycle progression, neoplastic cell transformation, and inflammation.[3,4,6–13] As an antiviral agent, catechins have exhibited the ability to inhibit reverse transcriptase and viral transcription, interfere with viral replication, and prevent adherence of viral particles to target cell receptors.[14,15,17–22]
To study the growth inhibitory potential of sinecatechins in HPV-positive tumor cell lines, four cervical carcinoma cell lines were treated with sinecatechins. The objectives of this study were to investigate growth inhibitory potential of the catechins in human cervical carcinoma cell lines infected with HPV. This includes investigating the impact of catechins on the viability of HPV-16 positive cells (CaSki and SiHa) and HPV-18 positive cells (HeLa and C4-I), assessing the kinetics of the antiproliferative effect of sinecatechins on cervical carcinoma cell lines, and determine whether the effect observed in the viability assays (WST-1) was due to a retardation in cell proliferation or due to reduction of total cell numbers leading to cell death. Furthermore, green tea extracts from different suppliers were evaluated to determine any differences in their growth inhibitory potential and further elucidate the potential MOA of sinecatechins in the eradication of EGW.
Methods
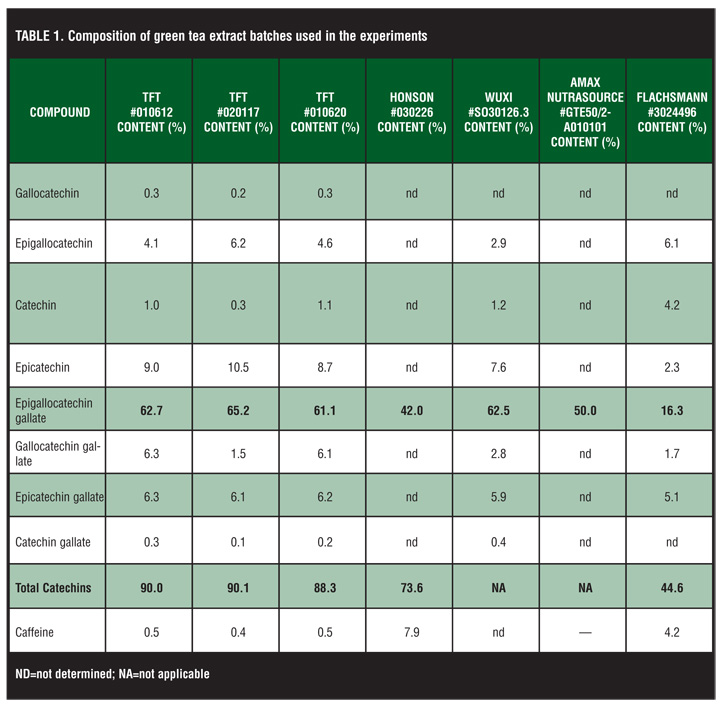
Three batches of green tea extracts from Tokyo Food Techno Co. Ltd., Tokyo, Japan, were used in the study. In addition, to compare different suppliers, green tea extracts from Honson Industries Ltd. (Markham, Canada), Wuxi (Wuxi Green Power BioProduct Ltd., Jiangsu Province, China), Amax NutraSource Inc (California, USA), and Emil Flachsmann AG (Wädenswil, Switzerland) were tested (for batch numbers and composition, see Table 1). The sinecatechins drug substance was synthesized from these green tea extracts. A molecular weight of 400g/mol was assumed for the calculations of sinecatechins and was dissolved in 0.9% NaCl to a concentration of 100mM. It was further diluted using the appropriate cell culture medium.
{kind=link}
Cultivation of cells. The human cervical carcinoma cell lines CaSki, SiHa (both carrying genomic HPV-16 copies), HeLa, and C4-I (both carrying genomic HPV-18 copies) were obtained from American Type Culture Collection (Manassas, Virginia, USA). CaSki cells were cultivated in RPMI 1640 containing 10mM HEPES-buffer, 2mM L-glutamine, 1mM sodium pyruvate, and 10% heat-inactivated fetal calf serum (FCS, Invitrogen, Karlsruhe, Germany). HeLa and SiHa cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 2mM L-glutamine, 1mM sodium pyruvate, 1% nonessential amino acids (Invitrogen). C4-I cells were cultured in Waymouth’s MB 752/1 medium supplemented with 10% heat-inactivated FCS (Invitrogen).
Viability assay (WST-1). CaSki, SiHa, HeLa, and C4-I cells were seeded in 96 well flat bottom plates. Four hours after seeding, cells were treated with increasing concentrations of sinecatechins. After two days, a WST-1 assay was performed as recommended by the manufacturer (Roche Diagnostics, Mannheim, Germany). This colorimetric viability assay quantifies cell viability and cell proliferation based on the cleavage of the tetrazolium salt WST-1 by mitochondrial dehydrogenases in viable cells. Cells were incubated with the WST-1 reagent for 4 to 6 hours. Then, the conversion of WST-1 was quantified in an enzyme-linked immunosorbent assay (ELISA)-reader at 450nm with a reference wavelength of 655nm. The amount of converted reagent is proportional to the amount of viable cells. The interassay variability was about 20 percent, whereas the intra-assay variability was up to 40 percent. The GI50 values were calculated by SigmaPlot 2004 for Windows Version 9.0 using a four-parameter sigmoid equation.
CaSki cells seeded in 96-well flat bottom plates (4.5×103 cells/well) were incubated with different concentrations of sinecatechins for increasing time periods. The medium was exchanged in cultures treated for less than 48 hours, that is, total incubation time was 48 hours in all samples. As a negative control, cells were incubated with only medium as described above. Viability was analyzed using the WST-1 assay. Cells were incubated with the WST-1 reagent for four hours. Samples were analyzed in triplicate.
For the analysis of different suppliers of green tea extract and different batches of sinecatechins, CaSki cells were seeded in a density of 4.5×103 cells/well in 96-well flat bottom plates. Green tea extracts were added and cells were cultivated for 48 hours. The viability was determined as described above using the WST-1 assay.
Clonogenic analysis. This assay was established to determine whether the effect observed in the viability assay (WST-1) was due to a retardation in cell proliferation or due to a reduction of total cell number leading to cell death. Therefore, HeLa and SiHa cells were seeded at low density (0.8×102 cells/well) in a six-well plate and cultivated for three days until small cell clones had developed. Cells were treated with sinecatechins in escalating concentrations (0.2mM–1mM) in medium containing 2.5% FCS for two hours. Thereafter, cells were cultivated in medium with 10% FCS for up to 23 days, until countable clones had developed. Medium exchange was carried out regularly.
Clones were stained with methylene blue (Merck, Darmstadt, Germany) for 10 minutes at room temperature. Then the plates were rinsed several times with distilled water and air-dried afterwards. The samples were tested in triplicate.
Results
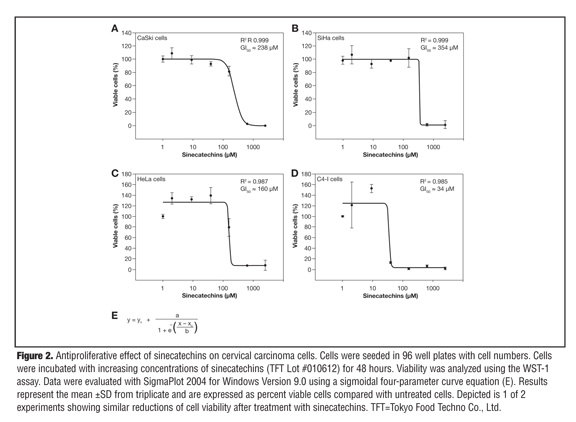
Antiproliferative effect of sinecatechins on cervical carcinoma cells. To analyze whether sinecatechins has an effect on the viability of HPV-16 and HPV-18 positive cells, two HPV-16 positive (CaSki, SiHa) and two HPV-18 positive cell lines (HeLa, C4-I) were tested. All four cervical carcinoma cell lines were incubated with increasing concentrations of sinecatechins (2–2500µM; TFT Lot. #010612) for 48 hours and thereafter the viability of cells was analyzed using a WST-1 assay. The cell growth of all four cell lines was inhibited in a concentration-dependent manner by sinecatechins (Figure 2). Total growth inhibition by sinecatechins was achieved at a concentration of 39µM for C4-I cells and of 625µM for CaSki, SiHa, and HeLa cells. With a calculated GI50 of approximately 34µM and a maximal inhibitory effect of 39µM, C4-I cells were most sensitive to the cytotoxic potential of sinecatechins (Figure 2D). CaSki, SiHa, and HeLa cells seemed to be less sensitive to the cytotoxic potential of sinecatechins than C4-I cells because their GI50 values ranged between 160µM and 360µM (Figure 2A-C). Interestingly, when cells were incubated with medium containing decreased amounts of FCS (2.5%), the growth inhibitory potential of sinecatechins was increased. These data demonstrate the concentration-dependent anti-proliferative effect of sinecatechins on HPV-16 and HPV-18 positive cervical carcinoma cell lines.
{kind=link}
Influence of the incubation time on the growth inhibitory effect of sinecatechins. To analyze the kinetic of the antiproliferative effect of sinecatechins on cervical carcinoma cell lines, CaSki cells were incubated with different concentrations of sinecatechins ranging from 78µM to 2.5mM for 1, 2, 4, 20, and 48 hours. Medium was exchanged in cultures treated for less than 48 hours (i.e., total incubation time was 48 hours in all samples). Viability was analyzed using the WST-1 assay. As negative control, cells were incubated with medium alone. In Figure 3, only two concentrations, 313µM and 625µM, are depicted because they were mostly affected by varying the treatment time with sinecatechins. Already a one-hour treatment was enough to detect a growth inhibitory effect at 313µM and 625µM (Figure 3). The analysis of the kinetic showed that the growth inhibitory potential increased with longer incubation times, reaching a maximum at 48 hours. The same type of experiment was performed using HeLa cells. The outcome was similar except for the fact that HeLa cells were more sensitive to treatment with sinecatechins, resulting in a 75-percent inhibition at 313µM after a four-hour treatment.
{kind=link}
Effect of green tea extract from different suppliers on the cell growth. To study whether green tea extracts from different suppliers have different growth inhibitory potential, three different batches from TFT (Tokyo Food Techno Co. Ltd., Tokyo, Japan) and one batch each from Honson Industries Ltd., Wuxi (Wuxi Green Power BioProduct Ltd), Amax NutraSource Inc, and Emil Flachsmann AG were tested (for composition, see Table 1). CaSki cells were treated with seven different concentrations of each batch ranging from 39µM to 2.5mM. Only 1 out of 7 batches showed a reduced potential to inhibit the growth of CaSki cells (Figure 4). EGCg is the major component of polyphenols in green tea extract. In the batches that showed similar growth inhibition potential, the EGCg content varied between 65 and 42 percent. Only in the sample from Flachsmann (#3024496) was the EGCg content (16%) and the total content of catechins (44.6%) drastically reduced. A reduced growth inhibitory effect was detectable in this sample as well.
{kind=link}
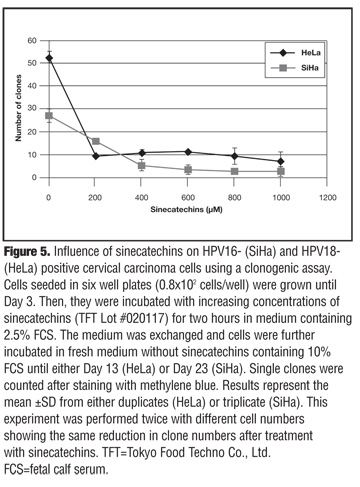
Influence of sinecatechins on cell growth using a clonogenic assay. To determine whether the effect observed in the viability assays (WST-1) was due to a retardation in cell proliferation or due to a reduction of total cell numbers leading to cell killing, a clonogenic assay was performed. HPV-16 and HPV-18 positive cervical carcinoma cell lines (SiHa and HeLa, respectively) were seeded in six-well plates and treated for two hours with increasing concentrations of sinecatechins. Then the medium was exchanged to remove sinecatechins and cells were further incubated until Day 17 and Day 23 for HeLa and SiHa cells, respectively.
The clone number revealed a reduction in both cell lines with increasing concentrations of sinecatechins (Figure 5). For HeLa cells, the highest drop in clone numbers was observed at 200µM, the lowest concentration of sinecatechins analyzed. The clone numbers dropped from about 50 to 10 clones. There was no further detectable decline in clone numbers using higher concentrations of sinecatechins (up to 1mM). In SiHa cells, treatment with sinecatechins resulted in a more constant decrease of clone numbers with increasing drug concentration.
{kind=link}
An almost total eradication of clones was obtained with 600µM of sinecatechins (Figure 5). The experiment was repeated with different cell numbers and the same effect was detected showing the long-term cytotoxic potential of sinecatechins in cervical cancer cell lines. Thus, sinecatechins do not only retard cell proliferation but also lead to cell death.
Discussion
To study the growth inhibitory potential of catechins in human cervical carcinoma cell lines infected with HPV, sinecatechins were studied against two HPV-16 positive (CaSki and SiHa) and two HPV-18 positive (HeLa and C4-I) cervical cell lines. Sinecatechins inhibited the cell growth of cervical cancer cell lines CaSki, SiHa, and HeLa by 50 percent at concentrations ranging between 160 to 360µM. Overall, the cell growth of all four cell lines was inhibited in a concentration-dependent manner by sinecatechins. C4-I cells were most sensitive to the treatment with sinecatechins, showing a GI50 of approximately 34µM, with maximum inhibitory effect of 39µM. Total inhibition was achieved at 625µM for CaSki, SiHa, and HeLa cells and 39µM for C4-I cells. The analysis of the growth inhibition kinetic showed that the effect increased with time, reaching a maximum at 48 hours. Furthermore, green tea extracts with an EGCg content ranging from 42 to 65 percent showed no differences in the growth inhibitory potential in a 48-hour assay. The batch from Flachsmann that contained only 16% EGCg did demonstrate drastically reduced growth inhibitory effect. The clonogenic assay confirmed the inhibitory effect on cell growth and showed that sinecatechins was not only able to inhibit the metabolic activity, but also led to cell death.
The range of concentrations of sinecatechins that affected the cell growth of tumor cell lines in this study is in concordance with findings reported by others.[4,12,28] Ahn et al[4] showed that EGCg inhibited the growth of CaSki cells with a GI50 of approximately 35µM in a 1- to 2-day assay. Yokohama et al[12] reported a 50-percent reduction of cell growth in SiHa cells at 100µM after a seven-day treatment (p<0.01) and a 25-percent inhibition at 100µM after four days in HeLa cells (p<0.05). Morré et al[28] showed a GI50 for the inhibition of HeLa growth of about 2µM in a 72-hour assay. The differences in the GI50 values may be due to differences in the incubation time, differences in the cell density, or differences in the assay used to detect growth inhibition. Interestingly, the study from Morré et al[28] showed that when individual green tea catechins (EGCg, GCg, ECg, EGC, and EC) were tested, EGCg was the most potent, followed by GCG. This is in concordance with the data from the current study, demonstrating that catechins containing a galloyl group are the most effective catechins in reducing cell viability. Morré et al[28] also reported that the amount of EGCg needed to inhibit cell growth was reduced about 10 times by combination of EGCg with the inactive catechin EC showing a synergistic effect between EGCg and EC.
The growth inhibitory potential of green tea extracts have been attributed to the induction of apoptosis, cell cycle arrest, and telomerase inhibition. In recent papers, the cytotoxic effect of green tea catechins in-vitro assays has also been said to be induced by increased levels of hydrogen peroxide (H2O2) in the cell culture medium. The production of ROS has been shown to be induced by some polyphenols during an auto-oxidation process under specific conditions in cell culture media. This antioxidant function plays a key role in enhancing the anti-inflammatory properties of EGCg, such as activation of transcription factors (NG-kB and AP-1) resulting in downregulation of inflammatory mediators. It is important to note that the impact of EGCg on metabolic activity that pronounces its antiproliferative effect and contributes to its overall cancer-preventative function may also make it possible for EGCg to direct antiviral activity.
Overall, this study shows the potential of sinecatechins to inhibit the growth of cervical cancer cells and this effect may in part contribute to its overall cancer-preventive function. The similarity of the growth inhibitory potentials of the various batches of green tea extract used to synthesize the sinecatechins drug substance may imply that the commercially available sinecatechins perform in the same manner. Although the exact MOA of sinecatechins is unknown, its known antioxidant, immunostimulatory, and antiproliferative activities, in addition to its antiviral effects may contribute to the high efficacy rates and low recurrence rates observed following its topical application for EGW treatment. Besides antioxidant activity, the FDA has not approved the inclusion of any additional activities in the product label. Most of the provider-administered and patient-applied EGW treatments rely on destructive methods to eradicate lesions, with the exception of imiquimod, which has immunostimulatory activity. No head-to-head trials with other topical patient-applied EGW therapies (i.e., imiquimod, podofilox) have been conducted; however, the EGW clearance rates reported in the published literature of the other treatments are lower than those observed following treatment with sinecatechins. More importantly, higher rates of recurrence have been reported (13–19% for imiquimod, 40% with cryotherapy, and as high as 91% with podofilox) compared with sinecatechins ointment, 15%.[27]
Conclusion
The inhibitory effects of green tea against experimental carcinogenesis and viral replication and inhibition have been demonstrated in vitro and/or in vivo in a variety of studies. As antineoplastic agents, catechins are described to inhibit cell growth and lead to apoptosis in several human carcinoma cells. However, as antiviral agents, catechins have been shown to inhibit viral replication through disruption during various stages of the viral life cycle, and prevent infectivity through processes, such as hemagglutination. Unlike anticarcinogenic processes, the antiviral ability of catechins has not been attributed to apoptosis or necrosis. The in-vitro activity of sinecatechins demonstrated in this study provides additional insight regarding its MOA as it relates to the clearance of EGW.
Acknowledgment
The author gratefully acknowledges MediGene AG, Planegg/Martinsried, Germany, and its employees for funding and conducting the study presented in this manuscript. The author also gratefully acknowledges the writing and editorial assistance of Priya Karkhanis and Malik Cobb, PA-C.
References
1. Cooper R, Morré DJ, Morré DM. Medicinal benefits of green tea: part I. Review of noncancer health benefits. J Altern Complement Med. 2005;11(3):521–528.
2. Meltzer SM, Monk BJ, Tewari KS. Green tea catechins for treatment of external genital warts. Am J Obstet Gynecol. 2009;200(3):233.e1–233.e7.
3. Balentine DA, Wiseman SA, Bouwens LC. The chemistry of tea flavonoids. Crit Rev Food Sci Nutr. 1997;37(8):693–704.
4. Ahn WS, Huh SW, Bae S-M, et al. A major constituent of green tea, EGCg, inhibits the growth of a human cervical cell line, CaSki cells, through apoptosis, G1 arrest, and regulation of gene expression. DNA Cell Biol. 2003;22(3):217–224.
5. Rahman I, Biswas SK, Kirkham PA. Regulation of inflammation and redox signaling by dietary polyphenols. Biochem Pharmacol. 2006;72(11):1439–1452.
6. Masuda M, Suzui M, Weinstein IB. Effects of epigallocatechin-3-gallate on growth, epidermal growth factor receptor signaling pathways, gene expression, and chemosensitivity in human head and neck squamous cell carcinoma cell lines. Clin Cancer Res. 2001;7(12):4220–4229.
7. Huh SW, Bae SM, Kim YW, et al. Anticancer effects of (-)-epigallocatechin-3-gallate on ovarian carcinoma cell lines. Gynecol Oncol. 2004;94(3):760–768.
8. Kuo PL, Lin C-C. Green tea constituent (-)-epigallocatechin-3-gallate inhibits Hep G2 cell proliferation and induces apoptosis through p53-dependent and Fas-mediated pathways. J Biomed Sci. 2003;10(2):219–227.
9. Gupta S, Ahmad N, Nieminen A-L, Mukhtar H. Growth inhibition, cell-cycle dysregulation, and induction of apoptosis by green tea constituent (-)-epigallocatechin-3-gallate in androgen-sensitive and androgen-insensitive human prostate carcinoma cells. Toxicol Appl Pharmacol. 2000;164(1):82–90.
10. Hastak K, Gupta S, Ahmad N, et al. Role of p53 and NF-kB in epigallocatechin-3-gallate-induced apoptosis of LNCaP cells. Oncogene. 2003;22:4851–4859.
11. Pianetti S, Guo S, Kavanagh KT, Sonenshein GE. Green tea polyphenol epigallocatechin-3 gallate inhibits Her-2/neu signaling, proliferation, and transformed phenotype of breast cancer cells. Cancer Res. 2002;62(3):652–655.
12. Yokoyama M, Noguchi M, Nakao Y, et al. The tea polyphenol, (-)-epigallocatechin gallate effects on growth, apoptosis, and telomerase activity in cervical cell lines. Gynecol Oncol. 2004;92(1):197–204.
13. Yang CS, Yang GY, Landau JM, et al. Tea and tea polyphenols inhibit cell hyperproliferation, lung tumorigenesis, and tumor progression. Exp Lung Res. 1998;24(4):629–639.
14. Chang L-K, Wei T-T, Chiu Y-F, et al. Inhibition of Epstein–Barr virus lytic cycle by (–)-epigallocatechin gallate. Biochem Biophys Res Commun. 2003;301(4):1062–1068.
15. Toda M, Okubo S, Ikigai H, et al. The protective activity of tea catechins against experimental infection by Vibrio cholerae O1. Microbiol Immunol. 1992;36(9):999–1001.
16. Weber JM, Ruzindana-Umunyana A, Imbeault L, Sircar S. Inhibition of adenovirus infection and adenain by green tea catechins. Antivir Res. 2003;58(2):167–173.
17. Yamaguchi K, Honda M, Ikigai H, et al. Inhibitory effects of (–)-epigallocatechin gallate on the life cycle of human immunodeficiency virus type 1 (HIV-1). Antivir Res. 2002;53(1):19–34.
18. Nakane H, Ono K. Differential inhibitory effects of some catechin derivatives on the activities of human immunodeficiency virus reverse transcriptase and cellular deoxyribonucleic and ribonucleic acid polymerases and various DNA and RNA polymerases by some catechin derivatives. Biochemistry. 1990;29:2841–2845.
19. Fassina G, Buffa A, Benelli R, et al. Polyphenolic antioxidant (–)-epigallocatechin-3-gallate from green tea as a candidate anti-HIV agent. AIDS. 2002;16(6):939–941.
20. Nakayama M, Suzuki K, Toda M, et al. Inhibition of the infectivity of influenza virus by tea polyphenols. Antivir Res. 1993;21(4):289–299.
21. Song J-M, Lee K-H, Seong B-L. Antiviral effect of catechins in green tea on influenza virus. Antiviral Res. 2005;68(2): 66–74.
22. Imanishi N, Tuji Y, Katada Y, et al. Additional inhibitory effect of tea extract on the growth of influenza A and B viruses in MDCK cells. Microbiol Immunol. 2002;46(7):491–494.
23. Okubo S, Toda M, Hara Y, Shimamura T. Antifungal and fungicidal activities of tea extract and catechin against Trichophyton. Fujieda, Japan; Mitsui Norin Co., Food Research Laboratories.
24. Mukoyama A, Ushijima H, Nishimura S, et al. Inhibition of rotavirus and enterovirus infections by tea extracts. Jpn J Med Sci Biol. 1991;44:181–186.
25. Yang CS, Maliakal P, Meng X. Inhibition of carcinogenesis by tea. Annu Rev Pharmacol Toxicol. 2002;42:25–54.
26. Veregen Product Monograph. Melville, NY: PharmaDerm, A Division of Nycomed US Inc.; 2010.
27. Tatti S, Stockfleth E, Beutner KR, et al. Polyphenon E®: a new treatment for external anogenital warts. Br J Dermatol. 2010;162:176–184.
28. Morré DJ, Morré DM, Sun H, et al. Tea catechin synergies in inhibition of cancer cell proliferation and of a cancer specific cell surface oxidase (ECTO-NOX). Pharmacol Toxicol. 2003;92(5):234–241.