David Weinstein, MD; Jennifer Leininger, MD; Carl Hamby, PhD; Bijan Safai, MD
Department of Dermatology, Department of Microbiology and Immunology, New York Medical College, New York and Valhalla, New York
Disclosure: The authors report no relevant conflicts of interest.
Abstract
Melanoma is a lethal melanocytic neoplasm. Unfortunately, the histological diagnosis can be difficult at times. Distinguishing ambiguous melanocytic neoplasms that are benign nevi from those that represent true melanoma is important both for treatment and prognosis. Diagnostic biomarkers currently used to assist in the diagnosis of melanoma are usually specific only for melanocytic neoplasms and not necessarily for their ability to metastasize. Traditional prognostic biomarkers include depth of invasion and mitotic count. Newer diagnostic and prognostic biomarkers utilize immunohistochemical staining as well as ribonucleic acid, micro-ribonucleic acid, and deoxyribonucleic acid assays and fluorescence in situ hybridization. Improved diagnostic and prognostic biomarkers are of increasing importance in the treatment of melanoma with the development of newer and more targeted therapies. Herein, the authors review many of the common as well as newer diagnostic and prognostic biomarkers used in melanoma.
(J Clin Aesthet Dermatol. 2014;7(6):13–24.)
Melanoma, an aggressive skin cancer, is currently the fifth most commonly diagnosed cancer in men and seventh in women in the United States with its incidence increasing 194 percent from 1975 to 2011.[1,2] In 2014, approximately 76,100 patients will be diagnosed with melanoma in the United States, accounting for an estimated 9,710 deaths.[1] Though recent advances in therapies for metastatic melanoma have shown some hope,[3,4] melanoma with distant metastasis still carries a grim prognosis with a five-year survival rate of 16 percent.[2] Given the poor prognosis for late stage melanoma, biomarkers are needed to aid in both the diagnosis and prognosis of melanoma and to determine which patients merit more aggressive therapy.
Diagnostic
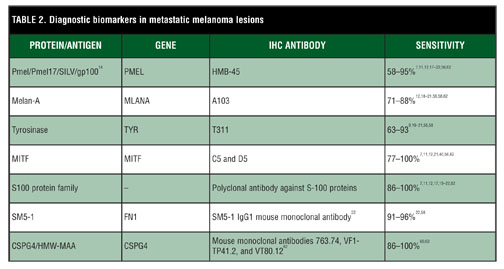
Immunohistochemical markers. The histological diagnosis of melanoma occasionally may be difficult due to its variety of cytomorphological variants. Melanoma can resemble different tumors, including carcinomas, neuroendocrine tumors, sarcomas, lymphomas, and germ cell tumors.[5] Therefore, immunohistochemical staining for melanocytic markers of differentiation often are employed in the diagnosis of melanonma.[6–12] Among the markers considered for use in the histological diagnosis of melanoma are Human Melanoma Black-45 (HMB-45), Melan-A, tyrosinase, microphthalmia transcription factor, and S100 as well as several newer ones (Table 1 and Table 2).
HMB-45 recognizes a 100 kD glycoprotein known as premelanosome protein (Pmel), Pmel17, gp100, or SILV.[13,14] Mutations in the Pmel gene result in a diluted, silver coat of normally black mice.[15] Pmel is found in pre-melanosomal vesicles and thought to be a necessary component of the fibrillar matrix for the polymerization of eumelanin.[14,16] HMB-45, a mouse monoclonal antibody, reacts with melanoma and junctional nevus cells.[17] Staining appears to be proportional to pigment content with lesions containing less pigment having little to no staining.[8] The sensitivity of HMB-45 has been shown to be 66 to 97 percent with decreased sensitivity in metastatic compared to primary lesions.[8,10,17–22] Specificity of distinguishing melanocytic from nonmelanocytic tumors is 91 to 100 percent.[10,19] Unfortunately, HMB-45 has demonstrated decreased specificity for malignant melanoma in sentinel lymph nodes compared to Melan-A.[23] As is common for many melanocytic biomarkers, HMB-45 demonstrates poor sensitivity for detecting desmoplastic malignant melanoma.[6,18,21,24]
Melan-A, also known as melanoma antigen recognized by T-cells-1(MART-1), is a melanocyte differentiation antigen expressed in the cytoplasm of both melanocytes, melanoma, and retinal pigmented epithelium.[25–27] It is a membrane protein located in melanosomes, endoplasmic reticulum, and the trans-Golgi network.[28] Melan-A associates with Pmel and is integral in its expression, trafficking, processing, and stability.[29] A recent study showed Melan-A to be superior to S–100 with a sensitivity of 93 percent and a specificity of 98 percent when differentiating melanoma and nonmelanocytic neoplasms.[10] However, other studies have shown lower sensitivities of 75 to 86 percent, with Melan-A less sensitive for detection of metastatic melanomas compared to primary melanomas.[18–21] Despite this, Melan-A has been found to be one of the most sensitive markers when used in frozen sections obtained during Mohs micrographic surgery.[30,31] While the specificity of Melan-A has been reported to be as high as 95 percent, a few concerns have been raised regarding its specificity.[19] Similar to HMB-45, Melan-A has lower sensitivity for desmoplastic melanoma.[18] Of note, it may be difficult to distinguish melanoma in situ from pigmented actinic keratoses and lichenoid reactions in sun-damaged skin based on Melan-A staining.[32,33] Melan-A also has been shown to stain adrenal cortical, Leydig, and granulosa and theca ovary cells as well as tumors derived from these cells.[18,19]
Tyrosinase, located in melanosomes, is an enzyme involved in the production of melanin.[34,35] Its sensitivity ranges from 90 to 100 percent for primary melanoma with decreasing sensitivity in later stage disease.[9,12,19–21] Specificity typically is very good at 97 to 100 percent when distinguishing melanoma from nonmelanocytic tumors.[19,21] As with other biomarkers, tyrosinase has reduced sensitivity in desmoplastic melanoma.[21]
Microphthalmia-associated transcription factor (MITF), a transcription factor of the MiT family, is a regulator of melanocyte development and differentiation and necessary for melanoblast differentiation from the neural crest.[36] Interestingly, as a transcription factor, MITF has been shown to regulate the transcription of Pmel, Melan-A, and tyrosinase.[37,38] Some early studies have demonstrated excellent sensitivity (100%), at times exceeding that of S-100 and HMB-45, and specificity for distinguishing melanoma from nonmelanocytic carcinomas.[7,11,39] However, later research has highlighted problems with the specificity of MITF due to its ability to stain histiocytes, lymphocytes, fibroblasts, Schwann cells and smooth muscle cells.[21,40] Some of these studies also showed a lower sensitivity of approximately 88 percent, though this was observed in metastatic lesions.[21] Furthermore, like other immunostains, MITF lacks sensitivity and specificity for desmoplastic or spindle cell melanomas.[21,24,39]
S-100, named for its 100 percent solubility in saturated ammonium sulfate, is a family of more than 24 proteins found in several different cell types, including glial cells, Schwann cells, melanocytes, Langerhans cells, and chondrocytes.[41–44] They exist both intracellulary as dimers and are secreted extracellularly. S-100 proteins are involved in many cellular functions including cell growth, cell cycle regulation, cell motility, calcium homeostasis, transcription, differentiation, regulation of cytoskeletal components and inflammatory responses among many others.[45–51] While its sensitivity is >89 percent in formalin fixed tissue,[8,10,19,20,22,52] S-100 staining may be less sensitive when used in frozen sections in Mohs micrographic surgery.[31] Despite high sensitivity, S-100 suffers from low specificity for melanoma, which is estimated to be 70 to 77 percent.[7, 10] This lack of specificity stems from the ability of S-100 to stain Schwann cells, chondrocytes, Langerhans cells, and myoepithelial cells among others as well as tumors derived from these cells.[22,53,54] Due to its lack of specificity, S-100 frequently is used simultaneously with more specific stains to distinguish melanoma from other S-100 positive malignancies. S-100 has much greater sensitivity compared to the above-mentioned biomarkers in desmoplastic malignant melanoma and thus, is of great utility in this variant of melanoma.[6,8,21,24,40,55,56]
SM5-1 is a new mouse IgG1 monoclonal antibody directed against two fibronectin isoforms that contribute to cell adhesion and migration and may play a role in melanoma metastasis.[57] In the few studies published so far, SM5-1 appears to be 95 to 99 percent sensitive for primary melanoma and 100 percent specific when distinguishing from other tumors tested; however, it is noted that it does stain perivascular dendritic cells, plasma cells, and myofibroblasts.[22,58] Like other markers, its sensitivity decreased in metastatic lesions, but only to 92 to 96 percent, which is much better than other currently available biomarkers.[22,58]
Chondroitin sulfate proteoglycan 4 (CSPG4), also known as high molecular weight melanoma-associated antigen (HMW-MAA) and melanoma chondroitin sulfate proteoglycan, is a membrane-bound proteoglycan found on melanocytes, endothelial cells, and pericytes among other cell types.[59] CSPG4 promotes cell adhesion, motility, and growth and may play a role in invasion and metastasis.[59]
Immunostaining with CSPG4 has demonstrated a sensitivity of >85 percent for melanoma with less sensitivity for benign melanocytic lesions, such as blue nevi.[60] While CSPG4 may have a lower sensitivity for acral lentignous melanoma, positive immunostaining has been associated with a worse prognosis for these lesions.[61] CSPG4 has a significant sensitivity of >90 percent for metastatic lesions, better than Melan-A, S-100, and HMB-45.[62] Desmoplastic melanoma frequently has diminished staining for many of the biomarkers; however, CSPG4 has recently been shown to have significantly greater sensitivity for detection of both primary and metastatic desmoplastic melanoma compared to HMB-45 and Melan-A.[63] CSPG4 also is interesting for its promising potential in immunotherapy for melanoma.[64,65]
A recent study showed that immunostaining of soluble adenylyl cyclase could assist in discriminating benign melanocytic nevi from melanoma. Specifically it was shown that the absence of the dot-like Golgi pattern and the presence of the pannuclear immunostaining was more indicative of melanoma.[66]
Immunohistochemical staining for p16 has recently been shown to greatly aid in distinguishing spitz nevi from melanoma. Decreased immunohistochemical staining of p16 has been shown to significantly correlate with the diagnosis of melanoma.[67-69]
In addition to the previously mentioned biomarkers used to aid in the diagnosis of melanoma, several other biomarkers currently are under investigation, including MUM-1, Mel-5, melanocortin-1, and PNL2 among others.[70,71]
While these biomarkers do facilitate the histopathological diagnosis of melanoma, Melan-A, HMB-45, and tyrosinase all show diminishing sensitivity with advancing stage disease.[12] Unfortunately, none of these biomarkers are able to distinguish malignant from nonmalignant melanocytic lesions.
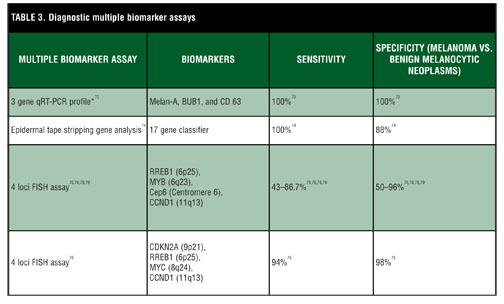
Biomarker panels and gene arrays. It is possible that several biomarkers together are needed to distinguish melanoma from melanocytic nevi. Furthermore, not every melanoma harbors the same mutations.[72] However, there may be several different mutations that a melanoma acquires in its progression toward cancer. Thus, particular combinations of mutations that result in upregulation or downregulation of certain biomarkers may be more consistent with or diagnostic of melanoma (Table 3). A study by Lewis et al73 illustrates this concept. Utilizing real-time quantitative reverse transcriptase-polymerase chain reaction,[73] they characterized the expression profile of 20 genes in melanoma, primary and metastatic, reactive lymph nodes, and benign nevi. Of the 20 genes utilized, three of them, Melan-A, budding uninhibited by benzimidazoles [1] homolog (BUB1), and CD [63], allowed for differentiation among melanoma, benign nevi, and lymphocytes. However, use of this set of genes was performed only on the training set from the study population and was not verified on a larger set of patients.
Another such example, an assay that analyzes multiple genes, has recently been shown to be both sensitive and specific in distinguishing melanoma from benign melanocytic nevi.[74] This innovative system utilizes tape stripping of melanocytic lesions to obtain corneocytes for ribonucleic acid (RNA) analysis in a noninvasive manner. The preliminary data from this study yielded a sensitivity of 100 percent and specificity of 88 percent for detection of melanoma or melanoma in situ though further clinical validation is needed.
Recent work by Gerami et al utilizing fluorescence in situ hybridization (FISH) assays showed promise in distinguishing ambiguous melanocytic tumors.[75 76] FISH has the benefit of being performed on paraffin-embedded tissue. Alhough the assay has demonstrated some use in discriminating benign from malignant melanocytic lesions, other studies have shown difficulty especially with spitzoid tumors.[77–80] Recent improvement of the assay utilizing the markers CDKN2A (9p21), RREB1 (6p25), MYC (8q24), and CCND1 (11q13) has shown increased sensitivity and specificity in addition to better discrimination of Spitz nevi from Spitzoid melanomas.[75] FISH analysis also can be of use adjunctively to distinguish lymph node nevi from melanoma metastasis.[81]
{kind=link}
{kind=link}
{kind=link}
Prognostic
Immunohistochemical biomarkers. While some patients will be cured with surgery alone, a significant number will not. Even patients with thin melanomas occasionally develop metastatic disease.[82] To help elucidate which patients are more likely to have disease progression and need adjuvant therapy, investigators have searched for histological prognostic biomarkers (Table 4).
While not considered a biomarker, the Breslow depth, or tumor thickness, on histopathology is the most accurate prognostic marker for patient survival in early stage cutaneous melanoma and hence, its inclusion in the American Joint Commission on Cancer (AJCC) melanoma staging system.[83,84]
Mitotic rate is currently included in the AJCC as one of the staging criteria because of its correlation with patient survival.[84] It is the second most significant predictor of patient survival, ranked behind only tumor thickness in localized primary cutaneous melanoma.[83]
Ki-67, a nuclear antigen, is a marker of proliferation that is expressed during the active phases of the cell cycle (G1, S, G2, and M).[85] For thin melanomas (<1mm), Ki-67 expression has been shown to correlate directly with prognosis and may correlate more highly with prognosis than mitotic count.[86,87] In addition, it has been shown that Ki-67 may be superior to mitotic count as a prognostic factor for survival in thicker melanomas (?1mm).[88] Furthermore, there is a high degree of interobserver variability among histopathologists in recognition of mitoses.[89]
Tumors with higher mitotic rates, Breslow thickness, and the absence of tumor infiltrating lymphocytes all are associated with an increased risk of sentinel lymph node involvement.[90] These markers take on even more importance since the number of nodal metastases is the single most significant predictor of patient survival in patients with stage III disease.[91]
BRAF mutations are found in more than 50 percent of melanomas, and of these, more than 90 percent consist of the V600E mutation.[92,93] These mutations can lead to constitutive activation of the MAPK pathway.[94] BRAF mutations, more specifically the V600E mutation, have not been associated with any significant difference in patient survival compared to those melanomas that lack this mutation.[95] However, the introduction of the BRAF inhibitor, vemurafenib, has been shown to improve survival in patients with late-stage melanoma that have the V600E mutation. Therefore, patients with tumors that are positive for this BRAF mutation may have improved survival with treatment due to this new treatment.[4]
Melanoma cell adhesion molecule (MCAM), also know as MUC18 and CD146, is a 113-kDa cell adhesion molecule normally expressed on endothelial and smooth muscle cells in adult tissue.[96] While rarely expressed in carcinomas, it is strongly expressed in advanced primary and metastatic melanoma and less so in nevi.[97–99] MCAM expression has been shown to be an independent predictor of prognosis in primary melanoma.[100, 101]
Metallothioneins are a family of heavy metal-binding low molecular weight proteins.[102] They contribute to the homeostasis of heavy metal ions and protect against oxidative stress as well as have several other roles.[102] Several studies have demonstrated that overexpression of metallothioneins in primary melanoma is associated with progression of disease and hematogenous metastasis.[103–105]
Recently, the biomarker CD10 has shown a significant correlation to progression and prognosis in patients with melanoma.[106–108] It has been proposed that CD10, a zinc-dependent endopeptidase, may affect prognosis by degradation of substances, such as enkephalin and substance P, which are known to suppress tumor progression in melanoma.[109,110] In one study, positive staining of CD10 correlated with a shorter five-year survival, although the majority of melanomas were acral lentiginous melanomas.[107]
Hundreds of studies investigating the myriad of molecular markers have been performed to better stratify patient risk and obtain improved prognostic information. Rothberg et al111 performed a systematic review of these studies and identified more than 100 proteins that represent potential candidates for prognostic markers in melanoma. However, many of these proteins had been evaluated only in a single study and not further substantiated.[111] Further research studies need to be conducted to evaluate their clinical utility as independent predictors of outcome in patients with melanoma.[111,112]
More recent research has utilized tissue microarrays to screen a wide panel of immunohistochemical markers and to conduct gene expression profiling to search for prognostic biomarkers and gene expression signatures.[72,113–118] As with the diagnostic biomarkers, it is unlikely that a single biomarker alone will be sufficient to determine prognosis. Perhaps a signature from an array or panel of biomarkers will most accurately predict prognosis. As such, several multimarker assays have recently been developed to more accurately predict prognosis (Table 5).
The combined score of the three biomarkers, NCOA3, SPP1, and RGS1, is significantly correlated with disease-specific survival and sentinel lymph node metastasis, making it an independent risk factor in primary cutaneous melanoma.[119] After evaluating 38 different markers, Rothberg et al120 recently designed a five-marker assay utilizing automated quantification of immunofluorescence that correlated significantly with reduced survival.[120]
Researchers likewise are investigating RNA, MicroRNA (miRNA), and deoxyribonucleic acid (DNA) assays, but those studies remain in the investigational phases of research. They currently are used to assist in selection of promising immunohistochemical biomarkers that may help determine prognosis and further elucidate the mechanisms of disease progression.[114–117]
RNA studies screening large numbers of genes have already revealed biomarkers that are significantly associated with prognosis. However, these studies are limited by the fact that they must be performed on cryopreserved tissue.[114–116] Nonetheless, recent studies have been able to ascertain profiles of RNA expression in paraffin-embedded, formalin-fixed melanoma tissue utilizing cDNA and new RNA extraction and isolation techniques.[117,121]
miRNA and DNA assays have the advantage of being conducted using paraffin-embedded, formalin-fixed tissue. Several studies utilizing miRNA already have shown that different profiles of miRNA are significantly associated with disease progression and survival.[107–110] Alhough fewer in number, DNA studies likewise have shown a correlation of different gene profiles with prognosis.[122]
FISH assays also may yield prognostic information. Positivity of one of the FISH assays already mentioned has been shown to be an independent risk factor for metastasis and melanoma-related death.[123] Another FISH assay evaluating copy number changes at CCND1 (11q13) and MYC(8q34) showed a gain of copy number in those melanomas that metastasized compared to those that did not.[124] Ideally, a complete genomic profile of the melanoma combined with the relevant prognostic information for each aberrantly expressed gene would give the most accurate prognosis, but the availability of such a prognostic indicator is still far off.
Serologic biomarkers. Serologic biomarkers have gained momentum in melanoma research in the search for the best markers of disease onset, progression, and therapeutic response. Using serologic markers is ideal as their testing is less invasive while providing a test with the potential to provide valuable information to clinicians treating melanoma patients. Melanoma-associated antigens, melanin-related metabolites, adhesion molecules, angiogenesis factors, and cytokines are among the serological biomarkers that are under current investigation.[125]
Lactate dehydrogenase (LDH), one of the earliest studied biomarkers in melanoma research, is a cytoplasmic enzyme responsible for the conversion of pyruvate to lactate. Cancer cells that replicate via anaerobic or glycolytic mechanisms have reduced dependence on oxygen for energy production creating a survival advantage.[126,127] This is relevant as tumors often have rapid growth resulting in necrosis and hypoxia as they quickly outgrow their vascular supply.[126] LDH elevations occur due to upregulation of LDH by tumor cells and by tumor cell necrosis causing spillover of the enzyme into the bloodstream.[126,127] In early melanoma research, elevated serum LDH levels were thought to be solely associated with liver metastasis; however, this has since been disproven.[128,129] Instead, elevated LDH levels have been consistently associated with adverse prognosis and directly correlate with survival in patients with stage IV disease.[84,129–131] Deichmann et al129 showed LDH to be the most specific biomarker for disease progression in stage IV melanoma patients with a 92 percent specificity though only 79 percent sensitivity.[129] False positives LDH in high-risk patients have been as high as 1.6 percent and can be due to hemolysis or other disease states, such as myocardial infarction.[132] LDH is the only current biomarker to be included in the AJCC 2009 staging system due to its significant prognostic value. This is evident in patients with stage IV disease and elevated LDH levels who have approximately 50 percent shorter one- and two-year survival rates compared to those patients with normal LDH levels.[84]
S100, used commonly as an immunohistochemical biomarker, as mentioned previously, also can be used as a serologic biomarker. While of limited value in early melanoma detection, elevated S100B levels have been found to be an indicator of advanced clinical disease stage.[133] Elevated S100B levels in advanced melanoma patients have been associated with metastasis, treatment response, relapse, and overall survival.[43,134–141] While LDH is a recognized important independent prognostic factor in advanced melanoma, it is primarily of value only in stage IV disease; S100 may be of equal if not superior value in monitoring and prognosis in stage III and IV disease.[132,141–144] S100B has been shown to have a false-positive rate of 1.9 percent and can also be elevated in cases of ischemic stroke, cerebrovascular disorders, and complications of cardio-bypass surgery.[132,133,145]
C-reactive protein (CRP) is a member of the pentraxin protein family that binds phosphocholine on bacteria and autologous ligands from necrotic and apoptotic cells and can activate complement.[146] As an acute phase reactant, it is a nonspecific marker of inflammation, infection, and tissue injury that is synthesized principally by hepatocytes in response to circulating cytokines including IL-6.[147–149] Elevated CRP has been associated with several malignancies in addition to a worse prognosis for those malignancies.[150] Since IL-6 levels correlate with tumor burden in melanoma, it is not surprising that IL-6 and, therefore CRP, correlate with disease progression.[151,152] Furthermore, increased CRP is associated with progression from stage I, II, or III to stage IV melanoma.[153] In patients receiving IL-2 immunotherapy, elevated CRP prior to initiating therapy was associated with a lack of response.[154]
Melanoma-inhibiting activity (MIA) is an 11 kd soluble protein, which has been characterized as an autocrine growth factor.[155] Despite the name MIA, hamster melanoma cells transfected with recombinant human MIA cDNA demonstrate increased invasiveness and metastasis of melanoma cells.[156] Higher levels of MIA are observed in melanoma compared to benign melanocytic nevi and normal skin.[157] Serum levels of MIA have been shown to correlate not only with disease stage, but also progression and response to therpay.[158–162] MIA is not specific for melanoma and can be elevated in other neoplasms, such as squamous cell carcinoma, late in pregnancy, and in children.[163,164]
Vascular endothelial growth factor (VEGF) is an angiogenic cytokine that regulates endothelial proliferation, differentiation, and survival.[165] Angiogenesis has been associated with solid-tumor growth, migration, and metastasis.[166,167] VEGF is secreted not only by melanoma cells, but also by peripheral blood lymphocytes and platelets, thus complicating its potential prognostic and clinical value.[168–170] While elevated VEGF is associated with disease stage, overall survival, progression of disease, and metastasis,[171,172] other studies have been less encouraging, showing elevation of VEGF when compared to controls, but no association with disease progression or therapeutic response.[173] In view of these limitations and the fact that VEGF has persistently shown lower sensitivity and specificity when compared to well-established biomarkers, its utility as a biomarker in melanoma is questionable.[173–175]
Reverse transcriptase-PCR (RT-PCR) and real-time quantitative PCR (qPCR) are techniques used to detect and quantify DNA and RNA expression, and have many applications within a wide variety of fields.[176] RT-PCR has been used on peripheral blood samples to assess for the presence of circulating melanoma cells through the detection of mRNA of the melanocyte specific gene tyrosinase.[177] Various melanoma markers in addition to tyrosinase, gp100, melan-A/MART1, MIA, p97, ?1è4-N-acetylgalactosaminyltransferase (GalNAc-T), paired box homeotic gene transcription factor 3 (PAX-3), and melanoma antigen A3 (MAGE-A3) have been used to detect circulating melanoma cells through both single and multimarker RT-PCR or qPCR.[177–181] While Palmieri et al showed that the presence of serum markers did not assist in prognosis, Arenberger et al found a rise in markers prior to disease progression and Koyanagi et al showed that the number of positive markers correlated with stage of disease.[178,179,181] Rarely, melanoma can metastasize across the placenta from mother to infant. qPCR was used successfully in a case to determine maternal tumor cell origin for proper diagnosis, prognosis, and management of the affected infant.[182]
Soluble BRAF V600E DNA mutations have been detected using RT-PCR and qPCR in patients with known cutaneous melanoma.[183,184] BRAF V600E, as mentioned previously, is currently used as a histological prognostic indicator and to guide available therapeutic options that target the BRAF-MEK-ERK pathway. Unfortunately, as a serological biomarker, BRAF V600E has not been useful in monitoring for disease progression. Among patients with melanoma, Pinzani et al184 found no correlation between serum BRAF V600E DNA levels and Breslow thickness, Clark level, presence of ulceration, nor sentinel lymph node positivity.[184]
Serum miRNAs are non-coding short RNA elements important in the regulation of gene expression and subsequent protein synthesis.[185] They help regulate cell proliferation, differentiation, and apoptosis and also affect expression of oncogenes and tumor suppressor genes.[186] Given the known dysregulation of miRNAs in cancer, they are currently being investigated for diagnostic, prognostic, and therapeutic utility.[187] Expression of miRNAs in tissue specimens already show correlation with diagnosis and prognosis.[188–191] Recently it has been discovered that miRNAs can be detected in the blood.[192] Circulating miRNAs show diagnostic and progonsotic utility in several different types of cancer, but have yet to be investigated in melanoma.[193]
Conclusion
Currently most diagnostic biomarkers of melanoma rely on detection of melanocytes rather than melanoma itself. Newer biomarkers depend on cytogenetic markers of carcinogenesis and signatures of mutations utilizing panels of biomarkers. There are no current serologic markers for the early detection of melanoma, and there may never be. Such evidence may be possible only in advanced stage disease that has metastasized from the primary site. As such, current serologic biomarkers detect circulating melanoma cells or secondary evidence of advanced disease, such as LDH.
Future research into serological and histological methods to detect early stages of melanoma hopefully will improve prognosis through earlier intervention. Such research might investigate markers of melanoma stem cells or markers of melanoblast differentiation that indicate a survival advantage and progression toward neoplasia.[194] Microarrays could be utilized to screen for similarities between melanoma and stem cells. Genes in common among melanoma and stem cells could be investigated to design better diagnostic and prognostic assays, perhaps using FISH, as well as to suggest new therapeutic targets.
{kind=link}
{kind=link}
References
1. The Surveillance, Epidemiology, and End Results (SEER) Program of the National Cancer Institute. SEER Stat Fact Sheets: Melanoma of the Skin. http://www.seer.cancer.gov/statfacts/html/melan.html. Accessed on May 26, 2014.
2. Howlader N, Noone AM, Krapcho M, et al (eds). SEER Cancer Statistics Review, 1975-2011, National Cancer Institute. Bethesda, MD. http://seer.cancer.gov/csr/1975_2011/, based on November 2013 SEER data submission, posted to the SEER website April 2014.
3. Robert C, Thomas L, Bondarenko I, et al. Ipilimumab plus dacarbazine for previously untreated metastatic melanoma. N Engl J Med. 2011;364(26):2517–2526.
4. Chapman PB, Hauschild A, Robert C, et al. Improved survival with vemurafenib in melanoma with BRAF V600E mutation. N Engl J Med. 2011;364(26):2507–2516.
5. Banerjee SS, Harris M. Morphological and immunophenotypic variations in malignant melanoma. Histopathology. 2000;36(5): 387–402.
6. Anstey A, Cerio R, Ramnarain N, et al. Desmoplastic malignant melanoma. An immunocytochemical study of 25 cases. Am J Dermatopathol. 1994;16(1):14–22.
7. Dorvault CC, Weilbaecher KN, Yee H, et al. Microphthalmia transcription factor: a sensitive and specific marker for malignant melanoma in cytologic specimens. Cancer. 2001;93(5):337–343.
8. Fernando SS, Johnson S, Bate J. Immunohistochemical analysis of cutaneous malignant melanoma: comparison of S-100 protein, HMB-45 monoclonal antibody and NKI/C3 monoclonal antibody. Pathology. 1994;26(1):16–19.
9. Hofbauer GF, Kamarashev J, Geertsen R, et al. Tyrosinase immunoreactivity in formalin-fixed, paraffin-embedded primary and metastatic melanoma: frequency and distribution. J Cutan Pathol. 1998;25(4):204–209.
10. Jing X, Michael CW, Theoharis CG. The use of immunocytochemical study in the cytologic diagnosis of melanoma: evaluation of three antibodies. Diagn Cytopathol. 2013;41(2):126–130. Epub 2011 Oct 24.
11. King R, Weilbaecher KN, McGill G, et al. Microphthalmia transcription factor. A sensitive and specific melanocyte marker for MelanomaDiagnosis. Am J Pathol. 1999;155(3):731–738.
12. Orchard GE. Comparison of immunohistochemical labelling of melanocyte differentiation antibodies melan-A, tyrosinase and HMB 45 with NKIC3 and S100 protein in the evaluation of benign naevi and malignant melanoma. Histochem J. 2000;32(8):475–481.
13. Adema GJ, de Boer AJ, Vogel AM, et al. Molecular characterization of the melanocyte lineage-specific antigen gp100. J Biol Chem. 1994;269(31):20126–20133.
14. Theos AC, Truschel ST, Raposo G, Marks MS. The Silver locus product Pmel17/gp100/Silv/ME20: controversial in name and in function. Pigment Cell Res. 2005;18(5):322–336.
15. Kwon BS, Halaban R, Ponnazhagan S, et al. Mouse silver mutation is caused by a single base insertion in the putative cytoplasmic domain of Pmel 17. Nucleic Acids Res. 1995;23(1):154–158.
16. Vennegoor C, Hageman P, Van Nouhuijs H, et al. A monoclonal antibody specific for cells of the melanocyte lineage. Am J Pathol. 1988;130(1):179–192.
17. Gown AM, Vogel AM, Hoak D, et al. Monoclonal antibodies specific for melanocytic tumors distinguish subpopulations of melanocytes. Am J Pathol. 1986;123(2):195–203.
18. Jungbluth AA, Busam KJ, Gerald WL, et al. A103: An anti-melan-a monoclonal antibody for the detection of malignant melanoma in paraffin-embedded tissues. Am J Surg Pathol. 1998;22(5):595–602.
19. Kaufmann O, Koch S, Burghardt J, et al. Tyrosinase, melan-A, and KBA62 as markers for the immunohistochemical identification of metastatic amelanotic melanomas on paraffin sections. Mod Pathol. 1998;11(8):740–746.
20. Gajjar NA, Cochran AJ, Binder SW. Is MAGE-1 expression in metastatic malignant melanomas really helpful? Am J Surg Pathol. 2004;28(7):883–888.
21. Miettinen M, Fernandez M, Franssila K, et al. Microphthalmia transcription factor in the immunohistochemical diagnosis of metastatic melanoma: comparison with four other melanoma markers. Am J Surg Pathol. 2001;25(2):205–211.
22. Trefzer U, Rietz N, Chen Y, et al. SM5-1: a new monoclonal antibody which is highly sensitive and specific for melanocytic lesions. Arch Dermatol Res. 2000;292(12):583–589.
23. Mahmood MN, Lee MW, Linden MD, et al. Diagnostic value of HMB-45 and anti-Melan A staining of sentinel lymph nodes with isolated positive cells. Mod Pathol. 2002;15(12):1288–1293.
24. Granter SR, Weilbaecher KN, Quigley C, et al. Microphthalmia transcription factor: not a sensitive or specific marker for the diagnosis of desmoplastic melanoma and spindle cell (non-desmoplastic) melanoma. Am J Dermatopathol. 2001;23(3): 185–189.
25. Chen YT, Stockert E, Jungbluth A, et al. Serological analysis of Melan-A(MART-1), a melanocyte-specific protein homogeneously expressed in human melanomas. Proc Natl Acad Sci U S A. 1996;93(12): 5915–5919.
26. Coulie PG, Brichard V, Van Pel A, et al. A new gene coding for a differentiation antigen recognized by autologous cytolytic T lymphocytes on HLA-A2 melanomas. J Exp Med. 1994;180(1): 35–42.
27. Kawakami Y, Eliyahu S, Delgado CH, et al. Identification of a human melanoma antigen recognized by tumor-infiltrating lymphocytes associated with in vivo tumor rejection. Proc Natl Acad Sci U S A. 1994;91(14):6458–6462.
28. De Maziere AM, Muehlethaler K, van Donselaar E, et al. The melanocytic protein Melan-A/MART-1 has a subcellular localization distinct from typical melanosomal proteins. Traffic. 2002;3(9): 678–693.
29. Hoashi T, Watabe H, Muller J, et al. MART-1 is required for the function of the melanosomal matrix protein PMEL17/GP100 and the maturation of melanosomes. J Biol Chem. 2005;280(14): 14006–14016.
30. Zalla MJ, Lim KK, Dicaudo DJ, Gagnot MM. Mohs micrographic excision of melanoma using immunostains. Dermatol Surg. 2000;26(8):771–784.
31. Albertini JG, Elston DM, Libow LF, et al. Mohs micrographic surgery for melanoma: a case series, a comparative study of immunostains, an informative case report, and a unique mapping technique. Dermatol Surg. 2002;28(8):656–665.
32. El Shabrawi-Caelen L, Kerl H, Cerroni L. Melan-A: not a helpful marker in distinction between melanoma in situ on sun-damaged skin and pigmented actinic keratosis. Am J Dermatopathol. 2004;26(5): 364–366.
33. Beltraminelli H, Shabrawi-Caelen LE, Kerl H, Cerroni L. Melan-a-positive “pseudomelanocytic nests”: a pitfall in the histopathologic and immunohistochemical diagnosis of pigmented lesions on sun-damaged skin. Am J Dermatopathol. 2009;31(3):305–308.
34. Raper HS. The Tyrosinase-tyrosine reaction: production from tyrosine of 5: 6-dihydroxyindole and 5: 6-dihydroxyindole-2-carboxylic acid-the precursors of melanin. Biochem J. 1927;21(1): 89–96.
35. Theos AC, Tenza D, Martina JA, et al. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Mol Biol Cell. 2005;16(11):5356–5372.
36. Hodgkinson CA, Moore KJ, Nakayama A, et al. Mutations at the mouse microphthalmia locus are associated with defects in a gene encoding a novel basic-helix-loop-helix-zipper protein. Cell. 1993;74(2):395–404.
37. Du J, Miller AJ, Widlund HR, et al. MLANA/MART1 and SILV/PMEL17/GP100 are transcriptionally regulated by MITF in melanocytes and melanoma. Am J Pathol. 2003;163(1):333–343.
38. Tachibana M, Takeda K, Nobukuni Y, et al. Ectopic expression of MITF, a gene for Waardenburg syndrome type 2, converts fibroblasts to cells with melanocyte characteristics. Nat Genet. 1996;14(1): 50–54.
39. King R, Googe PB, Weilbaecher KN, et al. Microphthalmia transcription factor expression in cutaneous benign, malignant melanocytic, and nonmelanocytic tumors. Am J Surg Pathol. 2001;25(1):51–57.
40. Busam KJ, Iversen K, Coplan KC, Jungbluth AA. Analysis of microphthalmia transcription factor expression in normal tissues and tumors, and comparison of its expression with S-100 protein, gp100, and tyrosinase in desmoplastic malignant melanoma. Am J Surg Pathol. 2001;25(2):197–204.
41. Cochran AJ, Lu HF, Li PX, et al. S-100 protein remains a practical marker for melanocytic and other tumours. Melanoma Res. 1993;3(5):325–330.
42. Donato R, Cannon BR, Sorci G, et al. Functions of s100 proteins. Curr Mol Med. 2013;13(1):24–57.
43. Schultz ES, Diepgen TL, Von Den Driesch P. Clinical and prognostic relevance of serum S-100 beta protein in malignant melanoma. Br J Dermatol. 1998;138(3):426–430.
44. Wen DR, Bhuta S, Herschman HR, et al. S-100 protein: a marker for melanocytic tumors. Ann N Y Acad Sci. 1983;420:261–266.
45. Donato R. Intracellular and extracellular roles of S100 proteins. Microsc Res Tech. 2003;60(6):540–551.
46. Heizmann CW, Fritz G, Schafer BW. S100 proteins: structure, functions and pathology. Front Biosci. 2002;7:d1356–1368.
47. Donato R. Functional roles of S100 proteins, calcium-binding proteins of the EF-hand type. Biochim Biophys Acta. 1999;1450(3): 191–231.
48. Donato R. S100: a multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int J Biochem Cell Biol. 2001;33(7):637–668.
49. Schafer BW, Heizmann CW. The S100 family of EF-hand calcium-binding proteins: functions and pathology. Trends Biochem Sci. 1996;21(4):134–140.
50. Zimmer DB, Cornwall EH, Landar A, Song W. The S100 protein family: history, function, and expression. Brain Res Bull. 1995;37(4):417–429.
51. Lin J, Yang Q, Wilder PT, et al. The calcium-binding protein S100B down-regulates p53 and apoptosis in malignant melanoma. J Biol Chem. 2010;285(35):27487–27498.
52. Clarkson KS, Sturdgess IC, Molyneux AJ. The usefulness of tyrosinase in the immunohistochemical assessment of melanocytic lesions: a comparison of the novel T311 antibody (anti-tyrosinase) with S-100, HMB45, and A103 (anti-melan-A). J Clin Pathol. 2001;54(3):196–200.
53. Nakajima T, Watanabe S, Sato Y, et al. An immunoperoxidase study of S-100 protein distribution in normal and neoplastic tissues. Am J Surg Pathol. 1982;6(8):715–727.
54. Kahn HJ, Marks A, Thom H, Baumal R. Role of antibody to S100 protein in diagnostic pathology. Am J Clin Pathol. 1983;79(3): 341–347.
55. Morgan MB, Purohit C, Anglin TR. Immunohistochemical distinction of cutaneous spindle cell carcinoma. Am J Dermatopathol. 2008;30(3):228–232.
56. Busam KJ, Kucukgol D, Sato E, et al. Immunohistochemical analysis of novel monoclonal antibody PNL2 and comparison with other melanocyte differentiation markers. Am J Surg Pathol. 2005;29(3): 400–406.
57. Trefzer U, Chen Y, Herberth G, et al. The monoclonal antibody SM5-1 recognizes a fibronectin variant which is widely expressed in melanoma. BMC Cancer. 2006;6:8.
58. Reinke S, Koniger P, Herberth G, et al. Differential expression of MART-1, tyrosinase, and SM5-1 in primary and metastatic melanoma. Am J Dermatopathol. 2005;27(5):401–406.
59. Campoli M, Ferrone S, Wang X. Functional and clinical relevance of chondroitin sulfate proteoglycan 4. Adv Cancer Res. 2010;109:73–121.
60. Natali PG, Giacomini P, Russo C, et al. Antigenic profile of human melanoma cells. Analysis with monoclonal antibodies to histocompatibility antigens and to melanoma-associated antigens. J Cutan Pathol. 1983;10(4):225–237.
61. Kageshita T, Kuriya N, Ono T, et al. Association of high molecular weight melanoma-associated antigen expression in primary acral lentiginous melanoma lesions with poor prognosis. Cancer Res. 1993;53(12):2830–2833.
62. Goto Y, Ferrone S, Arigami T, et al. Human high molecular weight-melanoma-associated antigen: utility for detection of metastatic melanoma in sentinel lymph nodes. Clin Cancer Res. 2008;14(11): 3401–3407.
63. Goto Y, Arigami T, Murali R, et al. High molecular weight-melanoma-associated antigen as a biomarker of desmoplastic melanoma. Pigment Cell Melanoma Res. 2010;23(1):137–140.
64. Schmidt P, Kopecky C, Hombach A. Eradication of melanomas by targeted elimination of a minor subset of tumor cells. Proc Natl Acad Sci U S A. 2011;108(6):2474–2479.
65. Torisu-Itakura H, Schoellhammer HF, Sim MS, et al. Redirected lysis of human melanoma cells by a MCSP/CD3-bispecific BiTE antibody that engages patient-derived T cells. J Immunother. 2011;34(8): 597–605.
66. Magro CM, Crowson AN, Desman G, Zippin JH. Soluble adenylyl cyclase antibody profile as a diagnostic adjunct in the assessment of pigmented lesions. Arch Dermatol. 2012;148(3):335–344. Epub 2011 Nov 21.
67. Al Dhaybi R, Agoumi M, Gagne I, et al. p16 expression: a marker of differentiation between childhood malignant melanomas and Spitz nevi. J Am Acad Dermatol. 2011;65(2):357–363.
68. Hilliard NJ, Krahl D, Sellheyer K. p16 expression differentiates between desmoplastic Spitz nevus and desmoplastic melanoma. J Cutan Pathol. 2009;36(7):753–759.
69. George E, Polissar NL, Wick M. Immunohistochemical evaluation of p16INK4A, E-cadherin, and cyclin D1 expression in melanoma and Spitz tumors. Am J Clin Pathol. 2010;133(3):370–379.
70. Ohsie SJ, Sarantopoulos GP, Cochran AJ, Binder SW. Immunohistochemical characteristics of melanoma. J Cutan Pathol. 2008;35(5):433–444.
71. Bhawan J. Mel-5: a novel antibody for differential diagnosis of epidermal pigmented lesions of the skin in paraffin-embedded sections. Melanoma Res. 1997;7(1):43–48.
72. Bittner M, Meltzer P, Chen Y, et al. Molecular classification of cutaneous malignant melanoma by gene expression profiling. Nature. 2000;406(6795):536–540.
73. Lewis TB, Robison JE, Bastien R, et al. Molecular classification of melanoma using real-time quantitative reverse transcriptase-polymerase chain reaction. Cancer. 2005;104(8):1678–1686.
74. Wachsman W, Morhenn V, Palmer T, et al. Noninvasive genomic detection of melanoma. Br J Dermatol. 2011;164(4):797–806.
75. Gerami P, Li G, Pouryazdanparast P, et al. A highly specific and discriminatory FISH assay for distinguishing between benign and malignant melanocytic neoplasms. Am J Surg Pathol. 2012;36(6): 808–817.
76. Gerami P, Jewell SS, Morrison LE, et al. Fluorescence in situ hybridization (FISH) as an ancillary diagnostic tool in the diagnosis of melanoma. Am J Surg Pathol. 2009;33(8):1146–1156.
77. Gerami P, Wass A, Mafee M, et al. Fluorescence in situ hybridization for distinguishing nevoid melanomas from mitotically active nevi. Am J Surg Pathol. 2009;33(12):1783–1788.
78. Vergier B, Prochazkova-Carlotti M, de la Fouchardiere A, et al. Fluorescence in situ hybridization, a diagnostic aid in ambiguous melanocytic tumors: European study of 113 cases. Mod Pathol. 2011;24(5):613–623.
79. Gaiser T, Kutzner H, Palmedo G, et al. Classifying ambiguous melanocytic lesions with FISH and correlation with clinical long-term follow up. Mod Pathol. 2010;23(3):413–419.
80. Raskin L, Ludgate M, Iyer RK, et al. Copy number variations and clinical outcome in atypical spitz tumors. Am J Surg Pathol. 2011;35(2):243–252.
81. Dalton SR, Gerami P, Kolaitis NA, et al. Use of fluorescence in situ hybridization (FISH) to distinguish intranodal nevus from metastatic melanoma. Am J Surg Pathol. 2012;34(2):231–237.
82. Gimotty PA, Elder DE, Fraker DL, et al. Identification of high-risk patients among those diagnosed with thin cutaneous melanomas. J Clin Oncol. 2007;25(9):1129–1134.
83. Thompson JF, Soong SJ, Balch CM, et al. Prognostic significance of mitotic rate in localized primary cutaneous melanoma: an analysis of patients in the multi-institutional American Joint Committee on Cancer melanoma staging database. J Clin Oncol. 2011;29(16): 2199–2205.
84. Balch CM, Gershenwald JE, Soong SJ, et al. Final version of 2009 AJCC melanoma staging and classification. J Clin Oncol. 2009;27(36):6199–6206.
85. Vereecken P, Laporte M, Heenen M. Significance of cell kinetic parameters in the prognosis of malignant melanoma: a review. J Cutan Pathol. 2007;34(2):139–145.
86. Gimotty PA, Van Belle P, Elder DE, et al. Biologic and prognostic significance of dermal Ki67 expression, mitoses, and tumorigenicity in thin invasive cutaneous melanoma. J Clin Oncol. 2005;23(31): 8048–8056.
87. Frahm SO, Schubert C, Parwaresch R, Rudolph P. High proliferative activity may predict early metastasis of thin melanomas. Hum Pathol. 2001;32(12):1376–1381.
88. Ladstein RG, Bachmann IM, Straume O, Akslen LA. Ki-67 expression is superior to mitotic count and novel proliferation markers PHH3, MCM4 and mitosin as a prognostic factor in thick cutaneous melanoma. BMC Cancer. 2010;10:140.
89. Barry M, Sinha SK, Leader MB, Kay EW. Poor agreement in recognition of abnormal mitoses: requirement for standardized and robust definitions. Histopathology. 2001;38(1):68–72.
90. Kruper LL, Spitz FR, Czerniecki BJ, et al. Predicting sentinel node status in AJCC stage I/II primary cutaneous melanoma. Cancer. 2006;107(10):2436–2445.
91. Balch CM, Gershenwald JE, Soong SJ, et al. Multivariate analysis of prognostic factors among 2,313 patients with stage III melanoma: comparison of nodal micrometastases versus macrometastases. J Clin Oncol. 2010;28(14):2452–2459.
92. Davies H, Bignell GR, Cox C, et al. Mutations of the BRAF gene in human cancer. Nature. 2002;417(6892):949–954.
93. Shinozaki M, Fujimoto A, Morton DL, Hoon DS. Incidence of BRAF oncogene mutation and clinical relevance for primary cutaneous melanomas. Clin Cancer Res. 2004;10(5):1753–1757.
94. Curtin JA, Fridlyand J, Kageshita T, et al. Distinct sets of genetic alterations in melanoma. N Engl J Med. 2005;353(20):2135–2147.
95. Akslen LA, Angelini S, Straume O, et al. BRAF and NRAS mutations are frequent in nodular melanoma but are not associated with tumor cell proliferation or patient survival. J Invest Dermatol. 2005;125(2):312–317.
96. Mintz-Weber CS, Johnson JP. Identification of the elements regulating the expression of the cell adhesion molecule MCAM/MUC18. Loss of AP-2 is not required for MCAM expression in melanoma cell lines. J Biol Chem. 2000;275(44): 34672–34680.
97. Bar-Eli M. Role of AP-2 in tumor growth and metastasis of human melanoma. Cancer Metastasis Rev. 1999;18(3):377–385.
98. Rummel MM, Sers C, Johnson JP. Phorbol ester and cyclic AMP-mediated regulation of the melanoma-associated cell adhesion molecule MUC18/MCAM. Cancer Res. 1996;56(9):2218–2223.
99. Shih LM, Hsu MY, Palazzo JP, Herlyn M. The cell-cell adhesion receptor Mel-CAM acts as a tumor suppressor in breast carcinoma. Am J Pathol. 1997;151(3):745–751.
100. Pearl RA, Pacifico MD, Richman PI, Wilson GD, Grover R. Stratification of patients by melanoma cell adhesion molecule (MCAM) expression on the basis of risk: implications for sentinel lymph node biopsy. J Plast Reconstr Aesthet Surg. 2008;61(3): 265–271.
101. Pacifico MD, Grover R, Richman PI, et al. Development of a tissue array for primary melanoma with long-term follow-up: discovering melanoma cell adhesion molecule as an important prognostic marker. Plast Reconstr Surg. 2005;115(2):367–375.
102. Thirumoorthy N, Shyam Sunder A, Manisenthil Kumar K, et al. A review of metallothionein isoforms and their role in pathophysiology. World J Surg Oncol. 2011;9:54.
103. Emri E, Egervari K, Varvolgyi T, et al. Correlation among metallothionein expression, intratumoural macrophage infiltration and the risk of metastasis inhuman cutaneous malignant melanoma. J Eur Acad Dermatol Venereol. 2012.
104. Weinlich G, Topar G, Eisendle K, et al. Comparison of metallothionein-overexpression with sentinel lymph node biopsy as prognostic factors in melanoma. J Eur Acad Dermatol Venereol. 2007;21(5):669–677.
105. Weinlich G, Eisendle K, Hassler E, et al. Metallothionein – overexpression as a highly significant prognostic factor in melanoma: a prospective study on 1270 patients. Br J Cancer. 2006;94(6): 835–841.
106. Bilalovic N, Sandstad B, Golouh R, et al. CD10 protein expression in tumor and stromal cells of malignant melanoma is associated with tumor progression. Mod Pathol. 2004;17(10):1251–1258.
107. Oba J, Nakahara T, Hayashida S, et al. Expression of CD10 predicts tumor progression and unfavorable prognosis in malignant melanoma. J Am Acad Dermatol. 2011;65(6): 1152–1160.
108. Bertucci F, Pages C, Finetti P, et al. Gene expression profiling of human melanoma cell lines with distinct metastatic potential identifies new progression markers. Anticancer Res. 2007;27(5A): 3441–3449.
109. Horvat S, Mlinaric-Majerski K, Glavas-Obrovac L, et al. Tumor-cell-targeted methionine-enkephalin analogues containing unnatural amino acids: design, synthesis, and in vitro antitumor activity. J Med Chem. 2006;49(11):3136–3142.
110. Manske JM, Hanson SE. Substance-P-mediated immunomodulation of tumor growth in a murine model. Neuroimmunomodulation. 2005;12(4):201–210.
111. Gould Rothberg BE, Bracken MB, Rimm DL. Tissue biomarkers for prognosis in cutaneous melanoma: a systematic review and meta-analysis. J Natl Cancer Inst. 2009;101(7):452–474.
112. Gould Rothberg BE, Rimm DL. Biomarkers: the useful and the not so useful–an assessment of molecular prognostic markers for cutaneous melanoma. J Invest Dermatol. 2010;130(8): 1971–1987.
113. Lin H, Wong RP, Martinka M, Li G. Loss of SNF5 expression correlates with poor patient survival in melanoma. Clin Cancer Res. 2009;15(20):6404–6411.
114. Haqq C, Nosrati M, Sudilovsky D, et al. The gene expression signatures of melanoma progression. Proc Natl Acad Sci U S A. 2005;102(17):6092–6097.
115. Winnepenninckx V, Lazar V, Michiels S, et al. Gene expression profiling of primary cutaneous melanoma and clinical outcome. J Natl Cancer Inst. 2006;98(7):472–482.
116. Journe F, Id Boufker H, Van Kempen L, et al. TYRP1 mRNA expression in melanoma metastases correlates with clinical outcome. Br J Cancer. 2011;105(11):1726–1732.
117. Conway C, Mitra A, Jewell R, et al. Gene expression profiling of paraffin-embedded primary melanoma using the DASL assay identifies increased osteopontin expression as predictive of reduced relapse-free survival. Clin Cancer Res. 2009;15(22): 6939–6946.
118. Alonso SR, Ortiz P, Pollan M, et al. Progression in cutaneous malignant melanoma is associated with distinct expression profiles: a tissue microarray-based study. Am J Pathol. 2004;164(1):193–203.
119. Kashani-Sabet M, Venna S, Nosrati M, et al. A multimarker prognostic assay for primary cutaneous melanoma. Clin Cancer Res. 2009;15(22):6987–6992.
120. Gould Rothberg BE, Berger AJ, Molinaro AM, et al. Melanoma prognostic model using tissue microarrays and genetic algorithms. J Clin Oncol. 2009;27(34):5772–5780.
121. Koh SS, Wei JP, Li X, et al. Differential gene expression profiling of primary cutaneous melanoma and sentinel lymph node metastases. Mod Pathol. 2012;25(6):828–837. Epub 2012 Mar 9.
122. Jonsson G, Busch C, Knappskog S, et al. Gene expression profiling-based identification of molecular subtypes in stage IV melanomas with different clinical outcome. Clin Cancer Res. 2010;16(13): 3356–3367.
123. North JP, Vetto JT, Murali R, et al. Assessment of copy number status of chromosomes 6 and 11 by FISH provides independent prognostic information in primary melanoma. Am J Surg Pathol. 2011;35(8): 1146–1150.
124. Gerami P, Jewell SS, Pouryazdanparast P, et al. Copy number gains in 11q13 and 8q24 [corrected] are highly linked to prognosis in cutaneous malignant melanoma. J Mol Diagn. 2011;13(3):352–358.
125. Brochez L, Naeyaert JM. Serological markers for melanoma. Br J Dermatol. 2000;143(2):256–268.
126. Agarwala SS, Keilholz U, Gilles E, et al. LDH correlation with survival in advanced melanoma from two large, randomised trials (Oblimersen GM301 and EORTC 18951). Eur J Cancer. 2009;45(10):1807–1814.
127. Koukourakis MI, Giatromanolaki A, Sivridis E, et al. Lactate dehydrogenase-5 (LDH-5) overexpression in non-small-cell lung cancer tissues is linked to tumour hypoxia, angiogenic factor production and poor prognosis. Br J Cancer. 2003;89(5): 877–885.
128. Finck SJ, Giuliano AE, Morton DL. LDH and melanoma. Cancer. 1983;51(5):840–843.
129. Deichmann M, Benner A, Bock M, et al. S100-Beta, melanoma-inhibiting activity, and lactate dehydrogenase discriminate progressive from nonprogressive American Joint Committee on Cancer stage IV melanoma. J Clin Oncol. Jun 1999;17(6): 1891–1896.
130. Sirott MN, Bajorin DF, Wong GY, et al. Prognostic factors in patients with metastatic malignant melanoma. A multivariate analysis. Cancer. 1993;72(10):3091–3098.
131. Weide B, Elsasser M, Buttner P, et al. Serum markers lactate dehydrogenase and S100B predict independently disease outcome in melanoma patients with distant metastasis. Br J Cancer. 2012;107(3):422–428.
132. Egberts F, Hitschler WN, Weichenthal M, Hauschild A. Prospective monitoring of adjuvant treatment in high-risk melanoma patients: lactate dehydrogenase and protein S-100B as indicators of relapse. Melanoma Res. 2009;19(1):31–35.
133. Kaskel P, Berking C, Sander S, et al. S-100 protein in peripheral blood: a marker for melanoma metastases: a prospective 2-center study of 570 patients with melanoma. J Am Acad Dermatol. 1999;41(6):962–969.
134. Bottoni U, Izzo P, Richetta A, et al. S100 serum level: a tumour marker for metastatic melanoma. Melanoma Res. 2003;13(4): 427–429.
135. Kruijff S, Bastiaannet E, Brouwers AH, et al. Use of S-100B to evaluate therapy effects during bevacizumab induction treatment in AJCC stage III melanoma. Ann Surg Oncol. 2012;19(2): 620–626.
136. Hauschild A, Engel G, Brenner W, et al. S100B protein detection in serum is a significant prognostic factor in metastatic melanoma. Oncology. 1999;56(4):338–344.
137. Martenson ED, Hansson LO, Nilsson B, et al. Serum S-100b protein as a prognostic marker in malignant cutaneous melanoma. J Clin Oncol. 2001;19(3):824–831.
138. Tarhini AA, Stuckert J, Lee S, et al. Prognostic significance of serum S100B protein in high-risk surgically resected melanoma patients participating in Intergroup Trial ECOG 1694. J Clin Oncol. 2009;27(1):38–44.
139. Bolander A, Agnarsdottir M, Wagenius G, et al. Serological and immunohistochemical analysis of S100 and new derivatives as markers for prognosis in patients with malignant melanoma. Melanoma Res. 2008;18(6):412–419.
140. Mocellin S, Zavagno G, Nitti D. The prognostic value of serum S100B in patients with cutaneous melanoma: a meta-analysis. Int J Cancer. 2008;123(10):2370–2376.
141. Kruijff S, Bastiaannet E, Kobold AC. S-100B concentrations predict disease-free survival in stage III melanoma patients. Ann Surg Oncol. 2009;16(12):3455–3462.
142. Banfalvi T, Boldizsar M, Gergye M, et al. Comparison of prognostic significance of serum 5-S-Cysteinyldopa, LDH and S-100B protein in Stage III-IV malignant melanoma. Pathol Oncol Res. 2002;8(3): 183–187.
143. Krahn G, Kaskel P, Sander S, et al. S100 beta is a more reliable tumor marker in peripheral blood for patients with newly occurred melanoma metastases compared with MIA, albumin and lactate-dehydrogenase. Anticancer Res. 2001;21(2B): 1311–1316.
144. Hamberg AP, Korse CM, Bonfrer JM, de Gast GC. Serum S100B is suitable for prediction and monitoring of response to chemoimmunotherapy in metastatic malignant melanoma. Melanoma Res. 2003;13(1):45–49.
145. Fagnart OC, Sindic CJ, Laterre C. Particle counting immunoassay of S100 protein in serum. Possible relevance in tumors and ischemic disorders of the central nervous system. Clin Chem. 1988;34(7): 1387–1391.
146. Thompson D, Pepys MB, Wood SP. The physiological structure of human C-reactive protein and its complex with phosphocholine. Structure. 1999;7(2):169–177.
147. Pepys MB, Hirschfield GM. C-reactive protein: a critical update. J Clin Invest. 2003;111(12):1805–1812.
148. Castell JV, Gomez-Lechon MJ, David M, et al. Interleukin-6 is the major regulator of acute phase protein synthesis in adult human hepatocytes. FEBS Lett. 1989;242(2):237–239.
149. Moshage HJ, Roelofs HM, van Pelt JF, et al. The effect of interleukin-1, interleukin-6 and its interrelationship on the synthesis of serum amyloid A and C-reactive protein in primary cultures of adult human hepatocytes. Biochem Biophys Res Commun. 1988;155(1): 112–117.
150. Allin KH, Nordestgaard BG. Elevated C-reactive protein in the diagnosis, prognosis, and cause of cancer. Crit Rev Clin Lab Sci. 2011;48(4):155–170.
151. Deichmann M, Benner A, Waldmann V, et al. Interleukin-6 and its surrogate C-reactive protein are useful serum markers for monitoring metastasized malignant melanoma. J Exp Clin Cancer Res. 2000;19(3):301–307.
152. Mouawad R, Benhammouda A, Rixe O, et al. Endogenous interleukin 6 levels in patients with metastatic malignant melanoma: correlation with tumor burden. Clin Cancer Res. 1996;2(8):1405–1409.
153. Deichmann M, Kahle B, Moser K, et al. Diagnosing melanoma patients entering American Joint Committee on Cancer stage IV, C-reactive protein in serum is superior to lactate dehydrogenase. Br J Cancer. 2004;91(4):699–702.
154. Tartour E, Blay JY, Dorval T, et al. Predictors of clinical response to interleukin-2–based immunotherapy in melanoma patients: a French multiinstitutional study. J Clin Oncol. 1996;14(5): 1697–1703.
155. Bogdahn U, Apfel R, Hahn M, et al. Autocrine tumor cell growth-inhibiting activities from human malignant melanoma. Cancer Res. 1989;49(19):5358–5363.
156. Guba M, Bosserhoff AK, Steinbauer M, et al. Overexpression of melanoma inhibitory activity (MIA) enhances extravasation and metastasis of A-mel 3 melanoma cells in vivo. Br J Cancer. 2000;83(9):1216–1222.
157. Bosserhoff AK, Hein R, Bogdahn U, Buettner R. Structure and promoter analysis of the gene encoding the human melanoma-inhibiting protein MIA. J Biol Chem. 1996;271(1):490–495.
158. Deichmann M, Benner A, Kuner N, et al. Are responses to therapy of metastasized malignant melanoma reflected by decreasing serum values of S100beta or melanoma inhibitory activity (MIA)? Melanoma Res. Jun 2001;11(3):291–296.
159. Cao MG, Auge JM, Molina R, et al. Melanoma inhibiting activity protein (MIA), beta-2 microglobulin and lactate dehydrogenase (LDH) in metastatic melanoma. Anticancer Res. 2007;27(1B): 595–599.
160. Bosserhoff AK, Kaufmann M, Kaluza B, et al. Melanoma-inhibiting activity, a novel serum marker for progression of malignant melanoma. Cancer Res. 1997;57(15):3149–3153.
161. Meral R, Duranyildiz D, Tas F, et al. Prognostic significance of melanoma inhibiting activity levels in malignant melanoma. Melanoma Res. 2001;11(6):627–632.
162. Stahlecker J, Gauger A, Bosserhoff A, et al. MIA as a reliable tumor marker in the serum of patients with malignant melanoma. Anticancer Res. 2000;20(6D):5041–5044.
163. Bosserhoff AK, Kuster H, Hein R. Elevated MIA levels in the serum of pregnant women and of children. Clin Exp Dermatol. 2004;29(6):628–629.
164. Sasahira T, Kirita T, Kurihara M, et al. MIA-dependent angiogenesis and lymphangiogenesis are closely associated with progression, nodal metastasis and poor prognosis in tongue squamous cell carcinoma. Eur J Cancer. 2010;46(12):2285–2294.
165. Connolly DT, Heuvelman DM, Nelson R, et al. Tumor vascular permeability factor stimulates endothelial cell growth and angiogenesis. J Clin Invest. 1989;84(5):1470–1478.
166. Folkman J. What is the evidence that tumors are angiogenesis dependent? J Natl Cancer Inst. 1990;82(1):4–6.
167. Redondo P, Sanchez-Carpintero I, Bauza A, et al. Immunologic escape and angiogenesis in human malignant melanoma. J Am Acad Dermatol. 2003;49(2):255–263.
168. Freeman MR, Schneck FX, Gagnon ML, et al. Peripheral blood T lymphocytes and lymphocytes infiltrating human cancers express vascular endothelial growth factor: a potential role for T cells in angiogenesis. Cancer Res. 1995;55(18):4140–4145.
169. Shellman YG, Park YL, Marr DG, et al. Release of vascular endothelial growth factor from a human melanoma cell line, WM35, is induced by hypoxia but not ultraviolet radiation and is potentiated by activated Ras mutation. J Invest Dermatol. 2003;121(4):910–917.
170. Wartiovaara U, Salven P, Mikkola H, et al. Peripheral blood platelets express VEGF-C and VEGF which are released during platelet activation. Thromb Haemost. 1998;80(1):171–175.
171. Ugurel S, Rappl G, Tilgen W, Reinhold U. Increased serum concentration of angiogenic factors in malignant melanoma patients correlates with tumor progression and survival. J Clin Oncol. 2001;19(2):577–583.
172. Redondo P, Bandres E, Solano T, Okroujnov I, Garcia-Foncillas J. Vascular endothelial growth factor (VEGF) and melanoma. N-acetylcysteine downregulates VEGF production in vitro. Cytokine. 2000;12(4):374–378.
173. Pelletier F, Bermont L, Puzenat E, et al. Circulating vascular endothelial growth factor in cutaneous malignant melanoma. Br J Dermatol. 2005;152(4):685–689.
174. Osella-Abate S, Quaglino P, Savoia P, et al. VEGF-165 serum levels and tyrosinase expression in melanoma patients: correlation with the clinical course. Melanoma Res. 2002;12(4):325–334.
175. Viac J, Schmitt D, Claudy A. Circulating vascular endothelial growth factor (VEGF) is not a prognostic indicator in malignant melanoma. Cancer Lett. 1998;125(1–2):35–38.
176. VanGuilder HD, Vrana KE, Freeman WM. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques. 2008;44(5):619–626.
177. Smith B, Selby P, Southgate J, et al. Detection of melanoma cells in peripheral blood by means of reverse transcriptase and polymerase chain reaction. Lancet. 1991;338(8777):1227–1229.
178. Arenberger P, Arenbergerova M, Vohradnikova O, Kremen J. Early detection of melanoma progression by quantitative real-time RT-PCR analysis for multiple melanoma markers. Keio J Med. 2008;57(1): 57–64.
179. Koyanagi K, Kuo C, Nakagawa T, et al. Multimarker quantitative real-time PCR detection of circulating melanoma cells in peripheral blood: relation to disease stage in melanoma patients. Clin Chem. 2005;51(6):981–988.
180. Mocellin S, Del Fiore P, Guarnieri L, et al. Molecular detection of circulating tumor cells is an independent prognostic factor in patients with high-risk cutaneous melanoma. Int J Cancer. 2004;111(5):741–745.
181. Palmieri G, Ascierto PA, Perrone F, et al. Prognostic value of circulating melanoma cells detected by reverse transcriptase-polymerase chain reaction. J Clin Oncol. 2003;21(5):767–773.
182. Raso A, Mascelli S, Nozza P, et al. Detection of transplacental melanoma metastasis using quantitative PCR. Diagn Mol Pathol. 2010;19(2):78–82.
183. Daniotti M, Vallacchi V, Rivoltini L, et al. Detection of mutated BRAFV600E variant in circulating DNA of stage III-IV melanoma patients. Int J Cancer. 2007;120(11):2439–2444.
184. Pinzani P, Salvianti F, Cascella R, et al. Allele specific Taqman-based real-time PCR assay to quantify circulating BRAFV600E mutated DNA in plasma of melanoma patients. Clin Chim Acta. 2010;411(17–18):1319–1324.
185. Esteller M. Non-coding RNAs in human disease. Nat Rev Genet. 2011;12(12):861–874.
186. Kasinski AL, Slack FJ. Epigenetics and genetics. MicroRNAs en route to the clinic: progress in validating and targeting microRNAs for cancer therapy. Nat Rev Cancer. 2011;11(12): 849–864.
187. Sandhu S, Garzon R. Potential applications of microRNAs in cancer diagnosis, prognosis, and treatment. Semin Oncol. 2011;38(6): 781–787.
188. Caramuta S, Egyhazi S, Rodolfo M, et al. MicroRNA expression profiles associated with mutational status and survival in malignant melanoma. J Invest Dermatol. 2010;130(8):2062–2070.
189. Luo C, Tetteh PW, Merz PR, et al. miR-137 Inhibits the Invasion of Melanoma Cells through Downregulation of Multiple Oncogenic Target Genes. J Invest Dermatol. 2013;133(3):768–775.
190. Mueller DW, Rehli M, Bosserhoff AK. miRNA expression profiling in melanocytes and melanoma cell lines reveals miRNAs associated with formation and progression of malignant melanoma. J Invest Dermatol. 2009;129(7):1740–1751.
191. Xu Y, Brenn T, Brown ER, Doherty V, Melton DW. Differential expression of microRNAs during melanoma progression: miR-200c, miR-205 and miR-211 are downregulated in melanoma and act as tumour suppressors. Br J Cancer. 2012;106(3):553–561.
192. Kosaka N, Iguchi H, Ochiya T. Circulating microRNA in body fluid: a new potential biomarker for cancer diagnosis and prognosis. Cancer Sci. 2010;101(10):2087–2092.
193. Palmer SR, Erickson LA, Ichetovkin I, et al. Circulating serologic and molecular biomarkers in malignant melanoma. Mayo Clin Proc. 2011;86(10):981–990.
194. Grichnik JM. Melanoma, nevogenesis, and stem cell biology. J Invest Dermatol. 2008;128(10):2365–2380.